蓝藻远红光吸收,机制是否是提高辐射产量的有效方案?
太阳光是地球上几乎所有生命能量的最终来源,而植物和藻类通过光合作将光能转化为化学能。近段时间,科学家们认为光合作的极限是700nm,波长超过700 nm的光无法驱动光合作用。然而,某些蓝藻可以突破这个限制,可以利用超过700 nm的远红光能量进行光合作用,他们是如何做到的呢?
远红光下的产氧光合作机制
光合作用是植物、藻类和蓝藻将光能转化为化学能的过程。光系统有两种类型分别为光系统I(PSI)和光系统II(PSII),核心依赖叶绿素a吸收红光(660 nm)和视觉(430 nm)的可见光,但对于700 nm以上的远红光几乎无效,理论上,其驱动水的氧化能量不低于1.8eV,但远红光的光子能量较低,仅为1.55eV,长期被认为无法支持产氧光合作用。
然而,总有例外,1996年,在澳大利亚海岸发现了一种蓝藻Acaryocholis marina,它几乎不依赖叶绿素a,而是用一种特殊的叶绿素d作为主要色素,可以吸收高达715 nm的远红光。这也是首次证明,光合作用的光谱范围可以突破传统界限。2010年,从Halomicronema hongdeloris 中分离出Chl f,可吸收709 nm的远红光。
叶绿素d和叶绿素f与叶绿素a的化学结构相似,但关键的差异在于它们多了一个甲酰基-CHO,这种改变让它们的吸收光谱向远红外偏移,2014年,研究发现某些蓝藻能在远红光下激活一套特殊的基因——FaRLiP基因簇,合成叶绿素d/f并重组整个光合系统,只有当蓝藻在远红光环境下时,才会启动一种特殊的适应机制。
远红光下的光合作用机制又是怎样的呢?光合系统如何捕获远红光,这些低能光子如何驱动水氧化和电子传递呢?光合系统中的能量传递依赖于色素间的距离、聚焦和能量级分配。光系统I和光系统II在远红光下的能量捕获效率不同。
光系统I通过红色形式(Red Forms)和低能色素漂移态高效捕获远红光,量子效率接近100%;而光系统II因反应中心效率较高,远红光效率较低,但某些蓝藻通过Chl d/f修饰反应中心提升效率。总的来说,光系统I的能级结构和色素排列更适合远红光,而PSII需通过反应中心改造实现低能光子利用。
远红光光合作需解决低能光子驱动电荷分离的挑战,光系统I叶绿素d和叶绿素f低能通过能电荷转移实现低能激发态的高效电荷分离,而光系统II叶绿素d/f修饰的反应中心通过提高氧化还原电位水氧化功能,补偿能量损失,但电子传递效率可能降低,导致部分能量无法利用。
2018年进行的一项关键实验,他们在仅提供>700 nm远红光的条件下培养蓝藻,结果发现这些蓝藻仍然能够进行光合作用,且光化学效率保持在传统光合生物的50%以上,相比之下,普通植物在相同的条件下几乎无法生存。
2018年进行的一项关键实验,他们在仅提供>700 nm的远红光的条件下培养蓝藻,结果发现这些蓝藻仍然能够进行光合作用,且光化学效率保持在传统光合生物的50%以上,相比之下,普通植物在相同的条件下几乎无法生存。
近期研究揭示了光合系统突破“红光极限”的分子策略,核心观点是光合生物通过色素化学修饰、蛋白质环境调节及反应中心重构,实现远红光的高效捕获与转化,但其效率与稳定性存在系统差异。
研究不仅深入探讨了对光合作用作物的理解,还为人工光合系统和作物光能效率的提升提供了理论检验机制。未来需结合合成生物学与生态学,探索这一应用边界,如水稻引入FaRLiP等,提高作物产量。
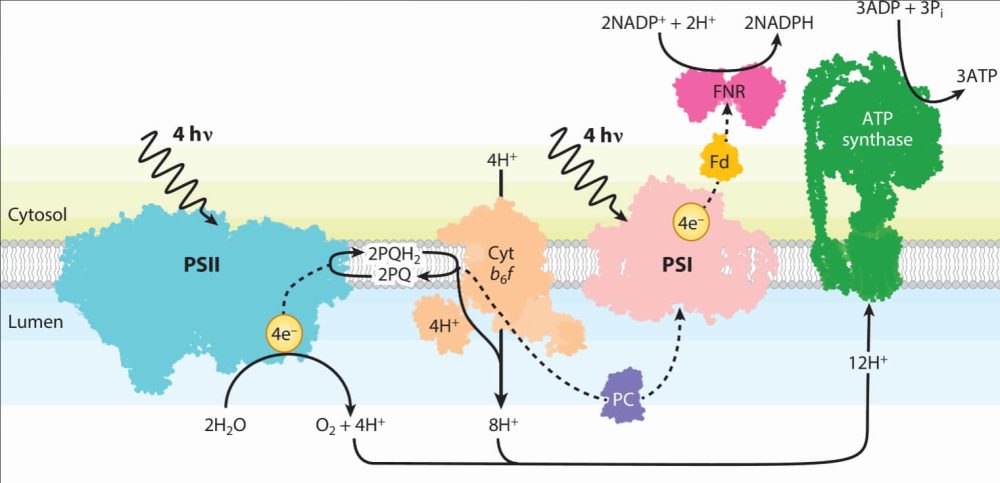
图:光合作用的光反应概述🔽
在所有产氧光合作用的生物体中,两种多色素复合物负责电子的转换。PSI和PSII复合物结合了丰富的色素(绿素和类胡萝卜素)。这些色素吸收光引起能转移到特定的叶绿素(称为反应中心),从而促进了分离。PSI和PSII位于类囊体膜内,它们通过细胞色素b 6f复合叶功能性连接,连接,从内部实现电子(PSII)把NADP +还原为NADPH(PSI)。电子转移过程中跨膜产生的质子逐渐被ATP合酶利用,产生ATP 。然后,NADPH和ATP在卡尔文-本森循环中被利用,将CO2固定为碳水化合物。在大多数生物体中,PSI和PSII 都具有天线系统,可以增加它们的吸收作用。虽然PSI和PSII在所有光合生物中高度警惕,但天线却并非如此。例如,在蓝藻中,天线通常由藻胆体组成,藻胆体是一种大型可溶性蛋白质组成体,使用胆素作为色素,位于膜的顶部。相反,在植物和绿藻中,天线系统主要由集光复合体多基因家族的成员组成,该家族由整合膜蛋白组成,并叶协调绿素和类胡萝卜素。不同类型的天线复合体存在于褐藻和硅藻以及相邻条件下的蓝藻中。缩写:cyt,细胞色素;PC,藻蓝蛋白;PSI/II,光系统I/II。
蓝藻如何突破光合作用极限?
据此,既然蓝藻能够如此高效地利用远红光,是否能够将这些机制转移到作物上呢?理论上,通过基因工程将叶绿素f合成酶引入作物,如水稻、小麦,让它们也能吸收远红光,但高等植物的光合系统结构比蓝藻复杂,如何让叶绿素f稳定整合到PSI和PSII中呢?
两种使蓝藻能够吸收远红光的兼性驯化过程分别为远红光光适应和弱光光驯化。远红光光适应通过缩小捕集光天线尺寸,增强光透明度,避免细胞的光保护性能量消耗散,缩小天线尺寸可提高培养体系整体光合效率。最近发现,弱光适应通过远红光吸收型藻蓝蛋白与PSI结合,扩大其吸收光谱范围,实现PSII与PSI的光谱分离,减少光竞争。
弱光光驯化解决了光合作用的效率低下问题,与 PSI 结合的吸收远红光的藻胆蛋白的表达,提供了一种分离每个光系统使用的光谱范围的机制。这可以通过优化来扩大氧光合作用中光利用的波长范围,并提高常用于生产生物燃料等有价值产品的蓝藻培养物的光合效率。
因此,自然光合作系统中的群体最新发现可能为提高光合作效率提供解决方案,这些改变对我们光合作用图片红光极限的发现,为利用合成生物学手段提高光合作效率提供了光明的前景。
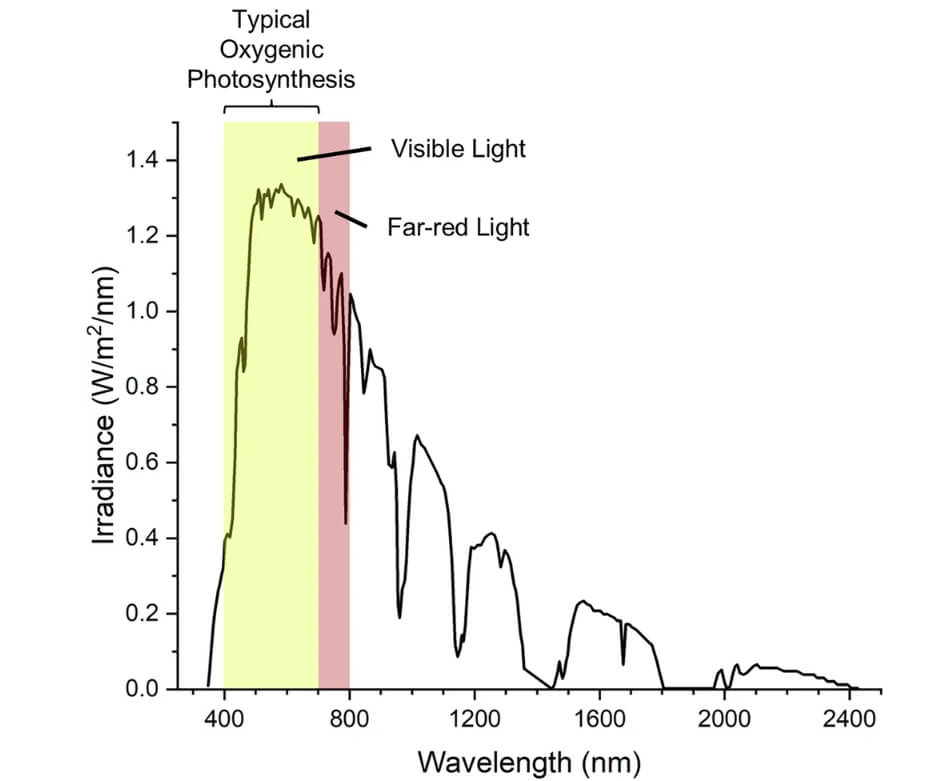
图:地球表面的太阳辐射光谱🔽
地球表面的太阳辐射光谱范围为250至2500纳米。图中突出显示了可见光和远红光区域。典型的产氧光合生物吸收可见光。
❓思考题:下列哪些是蓝藻突破光合作与“红光极限”的关键物质?
A.叶绿素a
B.叶绿素b/f
C.胡萝卜素
D.花青素
参考答案:(点击查看)
B.
解析:叶绿素d和叶绿素f与叶绿素a的化学结构相似,但关键的差异在于它们多了一个甲酰基-CHO,这种改变让它们的吸收光谱向远红外偏移。
参考文献:Christopher J. Gisriel & Gary W. Brudvig,Investigations into cyanobacterial photoacclimation processes address longstanding proposals for improving crop . 3942 ( 2025 ),doi:10.1038/s41467-025-59419-5